


Il mistero dell'Hobbitt. Un antenato in miniatura mette in crisi il darwinismo
di Mauro Quagliati - 30/01/2007
|
Apprendiamo dal National Geographic News del 27 ottobre 2004 che un nuovo membro è stato inserito nella variegata famiglia del genere umano, portando lo scompiglio tra i paleoantropologi: si tratta infatti niente di meno che di un “Hobbit”. O almeno così lo hanno battezzato affettuosamente i ricercatori indonesiani e australiani che ne hanno rinvenuto l’esemplare in una caverna dell’isola di Flores, a est di Bali. Si tratta di una serie di scheletri senza precedenti, che hanno già ricevuto l’onore di una nuova denominazione specifica: Homo floresienses. Con 1 metro di statura (per 25 kg), e circa 0,4 litri di capacità cranica, possedevano, da adulti, le proporzioni di un bambino di tre anni. Lo sconcerto cresce se si pensa che questi esseri hanno un’età giovanissima, compresa tra 18.000 e 15.000 anni.
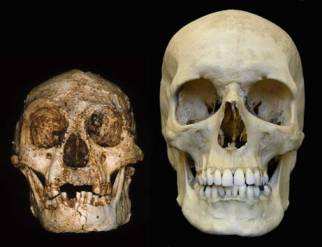
Figura 1. Confronto tra il cranio di Homo floresienses e uomo moderno. Peter Brown, tra i più noti paleoantropologi australiani, la considera una delle più spettacolari scoperte in mezzo secolo; l’omino di Flores è destinato a sollevare una quantità di problemi e di domande imbarazzanti poiché faticherà molto a trovare un posto logico sull’albero genealogico ufficiale dell’evoluzione umana.
Innanzitutto, questo reperto rappresenta la conferma definitiva che ominidi dalla morfologia anche molto diversa dalla nostra hanno convissuto con Homo sapiens praticamente fino all’altro ieri - in termini geologici - (in realtà l’Homo erectus di Giava era già stato post-datato a 30.000 anni, a metà degli anni ’90 [1]). Questo dovrebbe finalmente segnare l’abbandono definitivo del vecchio paradigma evoluzionista che prevedeva la successione/sostituzione graduale degli ominidi nella direzione evolutiva dell’uomo anatomicamente moderno. Al contrario, la nostra specie ha convissuto per decine di migliaia di anni con gli uomini di Neanderthal in Europa e con varianti di H. erectus nell’area australe.
Poi, fatto decisivo, demolisce completamente i criteri “volumetrici” (già lungamente criticati da molte parti) di distinzione tassonomica tra le specie ominidi. Pur avendo un cervello che occupa, in termini assoluti, un terzo del volume medio di H. sapiens (1250 cc) , gli antichi abitanti dell’Isola di Flores denotano statura eretta (che si evince da uno scheletro completo), hanno lasciato utensili, resti di fuoco e di cacciagione, per cui entrano a pieno titolo nel genere Homo. Sostanzialmente erano dei pigmei dalle caratteristiche estreme, ma con i tratti cranio-facciali tipici dell’H. erectus: arcate sopraorbitali sporgenti, cranio allungato e mento assente.
In terzo luogo si pone il quesito della provenienza e dell’origine filogenetica di questo gruppo (specie? sottospecie?). I ricercatori non possono far altro che postulare l’arrivo su Flores di una popolazione di H. erectus asiatico attorno a 800.000 anni fa, che si sarebbe sviluppato in miniatura, plausibilmente a causa di una pressione evolutiva “insulare” (si noti che sull’isola esisteva una specie di elefante nano, lo Stegodonte, estinto 12000 anni fa). Oppure si ipotizza che l’H. floresienses fosse già “pigmeo” prima di giungere sull’Isola. Ma questo non è cruciale, ciò che conta è che abbiamo un’ulteriore prova della grande plasticità delle popolazioni umane del pleistocene (1.700.000 - 12.000 anni fa), le cui caratteristiche fisiche suscitano, allo stato attuale della ricerca, una certa diatriba tra gli specialisti. Infatti è abituale considerare gli esemplari di quest’epoca, etichettati sotto le diverse denominazioni (Homo erectus, neanderthalensis, sapiens “arcaico”) come specie separate. Alcuni paleoantropologi invece le considerano adattamenti regionali di un’unica grande specie “politica” diffusa su tre continenti, nella quale, si può ora presumere, andrebbe ad inserirsi il pigmeo indonesiano. Semplificando il discorso, l’Homo pleistocenico ha dato vita a morfologie estreme rispetto a quelle dell’uomo anatomicamente moderno: il neandertaliano era più robusto e muscoloso degli Inuit attuali, l’Homo ergaster (Kenya, 1,7 milioni di anni) era più longilineo dei più alti Turkana odierni, e i “nani” di Flores erano più minuscoli del più basso pigmeo che si conosca.
Non basta, vi è un ulteriore problema che rende decisiva tutta la questione, ed è proprio la posizione geografica dell’Isola di Flores nell’arcipelago indonesiano.
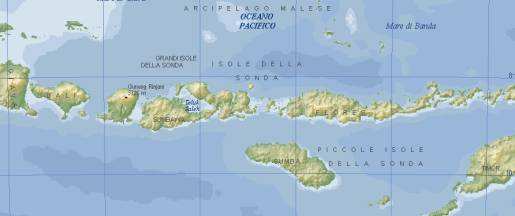
Fig. 2. L’isola di Flores, a 600 km a ovest di Giava, è separata da Bali da due bracci di mare profondo. Questa si trova isolata da ogni altra terra circostante da uno stretto di acqua profonda, denominato limite di Wallace, che separa la maggior parte della fauna asiatica da quella australiana. Chi colonizzò Flores nel passato doveva essere in grado di superare tale limite via mare, partendo da ovest (isola di Bali) superando due tratti di mare di una ventina di chilometri ciascuno, oppure da nord (Sulawesi) affrontando un viaggio ancora più lungo. Incredibile a dirsi, data la frammentarietà dei fossili che documentano le tappe evolutive dell’uomo, proprio questo sito era già diventato protagonista di una scoperta inaspettata, che si tinse dei toni gialli della cosiddetta “archeologia proibita”. Nel 1968, vennero rinvenuti sull’Isola degli utensili di pietra nello stesso strato degli stegodonti, di cui si conosceva l’età approssimativa di 750.000 anni. Ma, dato che l’autore della scoperta, il missionario olandese Theodor Verhoeven, non era un professionista del campo, la scoperta venne ampiamente trascurata. Una presenza di ominidi produttori di utensili era da considerarsi del tutto fuori luogo in un’isola sperduta oltre la barriera biologica di Wallace.
Figura 3. Utensili litici sull’isola di Flores datati a circa 800.000 anni. La datazione era assolutamente improponibile per una presenza umana (il quale, si sa, “deve” essere arrivato in Australasia non prima di 50.000 anni fa). Ed altrettanto problematica sarebbe stata l’attribuzione ad H. erectus, il quale, trenta anni fa, era ancora considerato un anello di congiunzione proto-scimmiesco, in grado sì di produrre utensili e, forse, comportamenti sociali rudimentali, ma ritenuto incapace di un’organizzazione e di una tecnologia sufficiente per affrontare il mare aperto, pur partendo dalla vicina Isola di Giava , in cui si trovava stanziato all’epoca.
Si è dovuto arrivare agli anni ’90 perché due misurazioni indipendenti, una paleomagnetica e una sulle ceneri vulcaniche, confermassero l’età dei reperti attorno a 840.000 anni. La situazione è così divenuta imbarazzante, con la comunità scientifica divisa tra gli estimatori delle inattese capacità di navigazione di H. erectus e chi invece tenta di minimizzare la scoperta.
In teoria si può ipotizzare l’emersione di ponti di terraferma dovuti dall’attività tettonica nell’area, in qualche momento del pleistocene, anche se fino ad oggi si suppone che in nessuna epoca geologica “recente” vi fosse un collegamento ininterrotto indo-australe: persino durante la massima escursione marina dell’ultima era glaciale quel tratto dell’arcipelago era coperto dal mare. Inoltre la fauna preistorica di Flores è composta da specie animali capaci di nuotare o, al limite, di andare alla deriva aggrappati a vegetazione galleggiante.
E’ stato anche suggerito che gli oggetti litici in questione non fossero effettivamente dei manufatti, insinuazione contro la quale Mike Morwood dell’Università australiana del New England è stato assolutamente categorico. Purtroppo quello di relegare potenziali utensili nella categoria dei “prodotti naturali”, quando questi vengano rinvenuti dove non dovrebbero stare, è un vizio secolare della paleoantropologia: con questa spiegazione di comodo, nella seconda metà dell’800, è passata sotto silenzio una solida evidenza della presenza umana nel pliocene e nel miocene europeo (vedasi Michael Cremo, Archeologia proibita, 1997).
Secondo i dati a disposizione sia i pigmei umani che la fauna pleistocenica caratteristica di Flores (oltre al suddetto stegodonte nano, la testuggine gigante e il varano gigante di Komodo) si sono estinti in seguito ad un’imponente eruzione vulcanica attorno a 12.000 anni fa. Anche se le evidenze archeologiche note di uomini moderni sulla nostra isola partono solo dal millennio successivo, è ora accertato che nell’area indo-australe vi sia stata una convivenza di almeno 20.000 anni tra uomini anatomicamente moderni e creature “nane”. Per cui l’ipotesi di relazioni culturali e di possibili incroci tra le diverse razze ha fatto subito capolino tra i ricercatori. Il tema delle ibridazioni tra il sapiens e i suoi predecessori pleistocenici è tuttora ampiamente dibattuta e, nonostante i dati genetici tendano a escludere ibridi tra le razze arcaiche e quelle moderne, vi è una vasta letteratura di comparazioni ossee e craniometriche che documentano la persistenza di caratteri ancestrali nelle popolazioni regionali dei diversi continenti. (in proposito si rimanda a “L’aborigeno australiano. un homo sapiens arcaico?” del sottoscritto). In quest’ottica, l’H. floresienses diventa allora un ritrovamento coerente, trovandosi proprio nel baricentro di un’area in cui sono insediati (o almeno sopravvivevano fino al secolo scorso) diverse popolazioni pigmee di colore, sparse su diverse isole dell’Oceano Indiano. Nel Golfo del Bengala, i “negritos” delle Isole Andaman presentano caratteri pigmoidi. Le zone montuose della penisola tailandese, malese e dell’Indonesia erano popolate, fino agli anni ’20, da etnie pigmee, oggi quasi completamente scomparse (i Semang della Malaysia, gli Yali dell’Indonesia).
Inoltre, fatto dimenticato dall’antropologia, anche in Australia, nel Queensland settentrionale, è stata ampiamente documentata la presenza di un’etnia pigmea. Le caratteristiche dei “Barrineans”, studiate da Norman B. Tindale and Joseph B. Birdsell negli anni’30, erano note agli antropologi e al vasto pubblico fino agli anni ’60. Si trattava di etnia di statura compresa tra 1,40 e 1,50 metri, con volti infantili, somiglianti agli estinti nativi della Tasmania (la popolazione più scura dell’Australia).
Pare che la memoria di questa popolazione, così come un’interessante teoria alternativa sull’origine delle popolazioni aborigene australiane, sia scomparsa dalla letteratura a partire dagli anni ’60, per motivi sostanzialmente politici. La teoria di Birdsell del “triplice ibrido” (trihybrid theory) suggeriva che i variegati tratti somatici delle numerose etnie aborigene (statura e corporatura, colore della pelle, tipo e colore del pelame) fossero il risultato di un rimescolamento di lungo periodo tra popolazione di origine, rispettivamente, pigmea, Vedda (chiari di pelle, con pelo folto caucasico, e tozzi) e negroide longilinea. Questa tesi, per lo meno suggestiva, è stata completamente censurata in favore dell’origine singola attraverso la migrazione africana recente. Il modello standard risultava infatti più funzionale alle rivendicazioni (sacrosante) del movimento per i diritti politici degli Aborigeni degli anni ’60, per il quale era opportuno unificare la lotta delle diverse etnie sotto un’unica bandiera, identificare cioè il diritto ancestrale alla terra sulla base della comune origine genetica. [2]
Eppure la scarsa popolarità dei pigmei isolani pare immotivata alla luce dell’origine africana recente. Non sarebbe forse un’ottima prova di una migrazione primitiva che partendo dal cuore tropicale-equatoriale dell’africa attraversò migliaia di chilometri toccando le coste e le isole dell’Indonesia fino alla Tasmania?
In realtà le cose non sono così semplici. Secondo le teorie ortodosse il primo uomo moderno (comparso tra 150.000 e 100.000 anni fa) dovrebbe essere un “normotipo” africano capace di adattarsi molto rapidamente ai diversi climi del mondo. Tanto rapidamente che la sua presenza è oggi attestata in Siberia, oltre il circolo polare artico già 40.000 anni fa.[3] La statura pigmea dovrebbe essere quindi un adattamento evolutivo secondario, abbastanza eccezionale, tipico di ambienti insulari e forestali (perché quindi la pelle scura?).
Trovare questi uomini in siti isolati e così distanti fra di loro difficilmente può essere imputato ad una improbabile riduzione corporea dei colonizzatori, intervenuta ripetutamente ad ogni successiva migrazione. Non è invece più logico ipotizzare un’antica stirpe umana originariamente distribuita su un ampio bacino tra Africa e Australasia? Non sarà casuale che le aree di sopravvivenza dei pigmei, siano zone marginali, distribuite a macchie di leopardo (montuose, forestali o completamente isolate come nel caso di Flores), come se queste etnie avessero già subito in passato una diaspora e una decimazione, probabilmente ad opera delle popolazioni che hanno colonizzato estensivamente il Pacifico.
Una prospettiva di questo tipo però implica la permanenza di tali caratteri fisici per molte generazioni, al di là di un estemporaneo “adattamento ambientale”, tanto da avvicinarla pericolosamente ad un concetto tabù della moderna antropologia, quello di “razza”. E’ risaputo che l’antropologia molecolare, ha minimizzato l’importanza delle differenze fisiche tra i tipi umani, per il fatto che il genoma sostanzialmente non le registra. Ma la fondamentale unità genetica della specie umana non è assolutamente incompatibile con il concetto di varietà umane: i “tipi”, in ogni specie, una volta manifestati, possono rimanere stabili per lunghi periodi, a meno di mescolamenti con altre razze interfeconde.
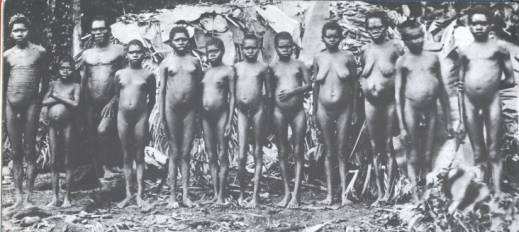
Proprio in relazione al pregiudizio razziale, possiamo trovare un altro dei motivi che hanno “cancellato” i pigmei dell’Oceania dalla memoria storica: si tratta di qualcosa che ha a che fare con la loro faccia. A guardare alcune foto d’epoca si notano frequentemente arcate sopraorbitali piuttosto sporgenti, indice di “primitività” quando si tratta di fossili, considerate una semplice ipertrofia ossea, quando invece si ha a che fare con uomini viventi. Purtroppo è noto il modo in cui questi caratteri sono stati fraintesi in senso razzista nel secolo scorso.
Figura 4. A sinistra Semang della Malesia. A destra tribù pigmea nei pressi di Cairns (1890). L’Indonesia rappresenta il nodo cruciale per il modello di popolamento dell’Oceania e qui si trovano una serie di ritrovamenti contraddittori che faticano sempre di più ad adattarsi alle idee classiche sulla migrazione degli esseri umani e ai preconcetti secondo cui il “gracile” e “moderno” deriva dal “robusto” e “primitivo”. I fossili conosciuti documentano la presenza di H. erectus in Asia a soli 300.000 anni dalla sua prima comparsa africana, la navigazione indonesiana di 800.000 anni fa, eppure non vi sono sue tracce nel continente australiano. Qui però stranamente sono stati trovati degli esseri umani recenti (10.000 anni) molto più robusti del normale [4]. E’ plausibile pensare che molti buchi nella serie fossile umana non siano dovuti solo all’aleatorietà della fossilizzazione, ma anche ad un processo di selezione semi-intenzionale dei reperti che, a quanto pare è ancora in corso. Man mano che ci si rende conto che l’uomo del pleistocene si comportava in maniera troppo umana per essere un gradino inferiore sulla scala evolutiva, potrebbe avvalorarsi la teoria della specie unica, secondo la quale i fossili degli ultimi 2 milioni di anni non sarebbero linee evolutive ramificate nella direzione di Homo sapiens, ma bensì adattamenti regionali di un’unica grande specie politipica, l’Uomo, nella quale includere i poli opposti, dal neandertaliano dei climi freddi al nuovo pigmeo dei tropici. Probabilmente i parametri di classificazione osteologici finora considerati validi per distinguere specie diverse dovrebbero essere riesaminati alla luce dell’estrema variabilità di questi antenati. Come si comporterebbe, si chiede la paleoantropologa Susan Anton, «un ricercatore che, tra un milione di anni, guardasse i pochi resti fossili di un pigmeo africano e di un giocatore dell’NBA»? [5]
Per concludere, va notato che questo episodio aggiunge un altro elemento ai già molti indizi, di carattere geologico e zoologico, che indicano nel periodo attorno al 10.000 a.C. la fine improvvisa di un equilibrio ecologico di lungo termine e probabilmente la fine della convivenza dell’uomo moderno con i membri più peculiari, forse specializzati, della famiglia. Alla luce di ciò si può suggerire che la denominazione convenzionale di “uomo anatomicamente moderno”, comprenda in realtà le razze fisicamente meno specializzate, che sono arrivate fino ad oggi.
Scampati alla grande crisi climatica della fine del pleistocene, sembra che i pigmei rimasti non sopravviveranno all’epoca moderna e alle politiche dei governi entro i cui confini sono capitati. Con la distruzione progressiva del loro ambiente nativo, l’omogeneizzazione culturale o il mescolamento con le popolazioni confinanti, in Africa come in Indonesia, questa antica Razza è destinata a lasciare la sua eredità solo sui libri di antropologia.
Torna alla pagina dei commenti
Per altri scritti di Mauro Quagliati vedi MMM GROUP (sito dedicato ad astroarcheologia e misteri).
NOTE 1. C. Swisher III et al., Latest Homo erectus of Java: Potential Contemporaneity with Homo sapiens in Southeast Asia, Science, 274 (1996). 2. In Nord America, argomentazioni simili hanno portato a sottostimare la diversità etnologica delle popolazioni native: purtroppo per una tragica ironia della storia, le rivendicazioni dei diritti dei Pellerossa americani hanno causato ostacoli allo studio del popolamento delle Americhe (come nel caso della diatriba sull’uomo di Kennewick). 3. P.Pavolov, J.I.Svendsen, S.Indrelid, Human presence in the European Arctic nearly 40,000 years ago, Nature, 2001. 4. Thorne, A. G., Macumber, P. G., Discoveries of Late Pleistocene man at Kow Swamp. Nature 238, 1972. 5. Susan Anton, National Geographic News, 25/3/2002. FONTI Hillary Mayell, "Hobbit" Discovered: Tiny Human Ancestor Found in Asia, National Geographic News 27/10/2004. Ancient mariners - Early humans were much smarter than we suspected, New Scientist, 14/3/1998 Ann Gibbons, Ancient Island Tools Suggest Homo erectus Was a Seafarer, Science 279,1998. Keith Windschuttle, Tim Gillin, The extinction of the Australian pygmies, Quadrant, June 2002. |